Rachel Merz
Walter Kemp Professor Emerita of Natural Science
Emeritus Biology
Contact
Affiliations: Biology
Research: The biomechanics of invertebrates

In my lab we examine ecological and evolutionary questions from a biomechanical perspective. We borrow theory and techniques from engineering and apply them to biological systems. The following are two recent examples:
(1) Testing Hypotheses About the Function of Polychaete Setae.
The polychaete annelids have existed for over 600 million years and have diversified to live in almost all marine ecosystems as well as some freshwater systems. In many habitats these worms play key roles in nutrient cycling and are important parts of food chains. Despite this, there are substantial gaps in our understanding of the biology of most species.

Polychaetes typically have chitinous hair-like structures called setae (=chaete) that extend from the body wall. These structures may be bundled together in tufts associated with the feet (parapodia) or in some cases they occur in rows or tori encircling the the body. Setae differ in their shape and sculpturing and in their distribution on the bodies of these worms.
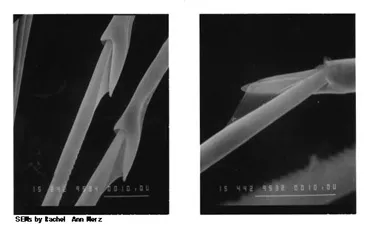
Figure 1. Scanning electron micrographs of the joints of compound or jointed setae of Ophiodromus pugettensis.

Figure 2. Scanning electron micrographs of the hooked setae of Sabellastarte magnifica.
Each species has a unique selection and distribution of setae. We want to know how the shapes of different setae and their distribution influences their function to the worms that bear them.
We have discovered that jointed setae help mobile worms move faster by providing better contact with the substratum.

Figure 3. Comparison of the change in the top locomotory speed for each gait before and after setal joints were removed. In the animals with intact joints there was no consistent pattern of change (diagonal striped lines). In the animals in which the joints were removed there was a general diminution of performance (solid black bars). (From: Merz, R. A. and D. R. Edwards 1998. Jointed-setae - their role in locomotion and gait transitions in polychaete worms. In Press, J. E. M. B. E.)
We have also demonstrated that for tube-dwelling polychaetes, hooked setae act as anchors.
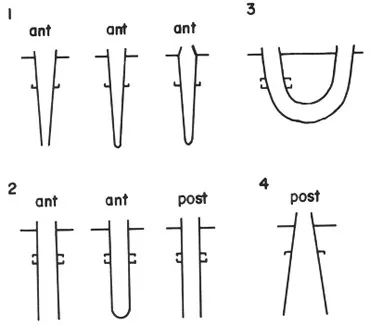
Figure 4. Predicted orientation of hooks relative to tube shape and position of openings. Solid horizontal line represents sediment surface. (From: Woodin, S. A. and R. A. Merz 1987. Holding on by their hooks: anchors for worms. Evolution 41(2): 427-432.)
(2) The Role of Flow in the Biology of Stream Dwelling Insects.
The life of stream dwelling insects is dominated by the movement of water around them. It brings food, removes wastes, distributes individuals and produces forces (lift and drag) that constantly act to reposition animals. To gain a better understanding of the specific ways in which flow acts we have studied the suspension feeding larvae of the black fly,Simulium vittatum. Our experiments are done in the lab in carefully designed flow tanks and at a variety of field sites.
We have been able to show that the posture of these insects larvae is dependent both on the velocity of the water and in the food concentration in the water.
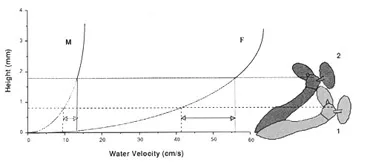
Figure 5. Model showing how an increase in water velocity yielded a change in the posture of a larval black fly depending on the velocity gradient in which it lives. (From: Hart, D. D., R. A. Merz, S. J. Genovese and B. D. Clark 1991. Feeding postures of suspension-feeding larval black flies: the conflicting demands of drag and food acquisition. Oecologia 85: 457-463.)
We have also have found that the black fly larvae can avoid one of their predators, the flatworm Dugesia dorotocephala, by positioning themselves in high flow environments where they also receive a benefit in increased feeding.
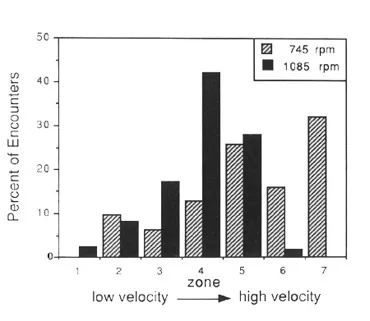
Figure 6. Distribution of encounters between larval black flies and predatory triclads depending on water velocity. In the slower treatment (745 rpm, triclads could attack larvae in all parts of the flow arena (zones 1-7). At higher speeds (1085 rpm), however, the triclads were virtually excluded from the areas of fastest flow (zones 6, 7). (From: Hansen, R. A., D. D. Hart and R. A. Merz 1991. Flow mediates predator-prey interactions between triclad flatworms and larval black flies. Oikos 60: 187-196.)
These results have been substantiated in the field, see: Hart, D. D. and R. A. Merz. 1998. Predator-prey interactions in a benthic stream community: a field test of flow-mediated refuges. Oecologia 114: 263-273.