|
|
|
Look Ma, No Archenteron!
Sulfate's role in sea urchin early development
Heather Sternshein,
Swarthmore College 2004
Adapted from "The effects of
sulfate on sea urchin development" Matthews Banda, F&M
2002
|
|
|
|
|

|
Objective
To observe the role sulfate plays in
sea urchin gastrulation, and to replicate the findings of
Karp and Solursh, that sea urchin embryos fail to gastrulate
without sulfate. To test whether endoderm differentiation
can occur in the absense of the movements of
gastrulation.
Abstract
Sea urchin development requires not only internally
produced signals, but also materials incorporated from the
environment. Karp and Solursh have hypothesized that
secondary mesenchyme cells, which form the filopodia of the
developing archenteron (primary gut) require sulfate (to
form sulfated proteoglycans which act as something like an
adhesive) in order to migrate along the acid
mucopolysaccharide of the extracellular matrix within the
blastocoel of a developing sea urchin (1974). Developing sea
urchin embryos were raised in either artificial sea water or
sulfate-free sea water, fixed, and stained for alkaline
phosphotase enzyme activity and with immunofluorescent
antibodies to vegetal archenteron cells. Embryos raised in a
sulfate-free environment, unlike controls, did not develop
archenterons. Staining indicated, however, that these cells
did differentiate to become gut cells and had invaginated,
but the cells could not migrate up the blastocoel wall. This
data supports the findings and hypothesis of Karp and
Solursh (1974).
Introduction
Sea urchin development progresses in a
predictable and easily observable way (Gilbert, 2000). The
vegetal plate thickens and primary mesenchyme cells ingress
and form spicules, the urchin skeleton (Figure 1). The
vegetal plate then invaginates, forming the archenteron, or
primitive gut. The archenteron migrates up the sea urchin's
blastocoel wall with the help of secondary mesenchyme cells
(Figure 1).
The migration of the archenteron
depends not only on signals and proteins already present in
the egg, but also on extracellular materials that have been
incorporated into the organism. Karp and Solursh have
hypothesized that secondary mesenchyme cells, which form the
filopodia of the developing archenteron (primary gut)
require sulfate (as something like an adhesive) in order to
migrate along the extracellular matrix within the blastocoel
of a developing sea urchin (1974).
Presumably, sea urchin embryos
incorporate sulfate from the environment into their
extracellular matrixes. The extracellular matrix contains
acid mucopolysaccharide, which when bound to sulfate, is
rough in appearance (Karp and Solursh, 1974). This roughness
is akin to velcro's roughness, allowing secondary mesenchyme
cells to pull the archenteron up along the blastocoel
cavity. If sulfate is not present, it has been observed that
an archenteron does not form (Karp and Solursh, 1974). Fixed
and stained embryos will indicate if the gut endoderm cells
have differentiated (and have simply failed to
migrate).
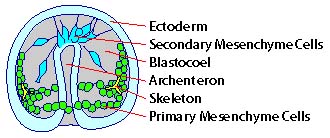
Figure 1. A schematic drawing of a
gastrulating sea urchin embryo. Note the secondary
mesenchyme cells, in the form of a filopodia, attaching to
the blastocoel wall to pull the archenteron up through the
blastocoel to form the gut cavity.
|
|
