|
|
|
|
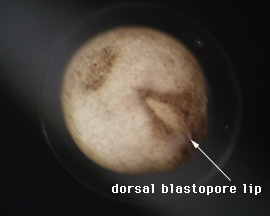
DEVELOPMENTAL EFFECTS OF LITHIUM CHLORIDE ON XENOPUS EMBRYOS Justin A Spenillo, Franklin & Marshall College PURPOSE: The objective of this experiment is to study the effect of lithium chloride treatment on early blastula xenopus embryos. INTRODUCTION: When the developing embryos of many organisms, such as the lytechinus variegatus (sea urchin) and xenopus (frog), are treated with lithium chloride, they experience alterations or perturbations in development. Lithium is believed to be the cause these perturbations due to its ability to inhibit glycogen synthase kinase-ß(beta) which initiates developmental pathways. The earliest stages of Xenopus development after fertilization of the egg is characterized by the rapid cleavage of cells, which is holoblastic(uneven) due to the large concentration of yolk at the vegetal pole. After the blastula has formed and the blastocoel, the hollow cavity inside the blastula, is present, then gastrulation begins. Gastrulation initiates on the future dorsal side of the organism which is marked by the gray crescent. The gray crescent is the region which is opposite from the location of sperm entry. This region has rotated, the cortical(outer) cytoplasm rotates 30ű, so that its location is between the animal and vegetal pole. The cells which are migrating into the embryo travel in through the dorsal blastopore lip. The dorsal blastopore lip is not only the point of entry of cells, but also sets the dorsal-ventral axis and marks the posterior of the embryo. When early embryos are treated with lithium chloride before the 64 cell stage, it has been shown that the embryos undergo dorsalization (Kao and Elison 1988, 1989; Slack 1988). Dorsalization is when the dorsal blastopore lip forms around the entire embryo at the same time. The result is an embryo that has a continous doral side. The most extreme results of this treatment are embryos with radial symmetric heads, a cylindrical heart, and bands of eye pigment. (Sive et al. 2000) This event occurs at the same time in development that the dorsal blastopore lip would form if there had been no lithium chloride treatment administered. As mentioned, lithium has the ability to inhibit glycogen synthase kinase-ß(beta), which is a substance which initites the wnt pathway in xenopus embryos that leads to dorsal axis formation (Klein and Melton 1996). PROTOCOL: 1. Dejelly early blastula embryos before the 32 cell stage.
2. Rinse healthy embryos and transfer 3/4 to the experimental petri dish containing 0.3M lithium chloride in 0.1x MBS. Transfer the remaining embryos to a control petri dish with 10% Steinberg's solution. 3. Gently mix solution. 4. Incubate embryos at 18-20űC for 10 minutes. 5. Rinse incubated embryos and transfer to petri dish with 10% Steinberg's solution. 6. Incubate embryos at 18-20űC and score perturbations at early gastrula. * Protocol adapted from Early Development of Xenopus Laevis.
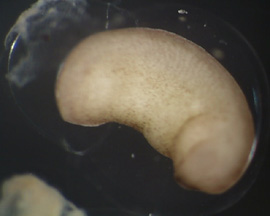
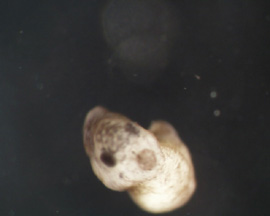
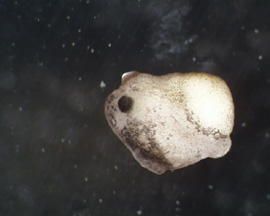
RESULTS: After 1 day, the control emryos were developing at a normal pace and were complete with gastrulation. As Figure 1 and 2 show, the embryos were far along in development and nearly at the tadpole stage. Lithium exposed embryos were much clower in develoment at the one day stage. At the one day mark, the dorsal blastopore lip had formed but was irregular and elongated; it had the appearance of an "eye" as can be seen in Figure 3 and 4. This follows Kao and Elison's work which says that lithium will cause a pertubation, called dorsalization, of the dorsal blastopore lip.
RESULTS(continued): After 2 days, the control group has developed into normal, functioning tadpoles (Figure 11 - 12). The lithium exposed group is undergoing a nontraditional developmental pattern. As can be seen in Figure 5 - 8, the embryos have no defined anterior except for the black dot which is the mouth. The organisms as a whole have minimal anterior-posterior definition and appear to be developing radially, all of tissues are forming in multiple numbers as a result of the dorsalization of the blastopore lip. In Figure 9 - 10, the embryos appear to have developed to unhealthy tadpoles; this is unlikley to have been caused by the lithium. None of the lithium exposed embryos survived past the 2 day stage; they disintegrated.
CONCLUSIONS: This experiment successful exhibited the ability of lithium to alter the development of the frog embryos. The control organism developed normally, while the lithium induced embryos underwent dorsalization, altered development, and death as predicted by previous research. The altered development of the embryos was marked by a minimal anterior-posterior and dorsal-ventral axis. If the embryos survived longer, radially development would most llikley have been exhibited. Lithium has been shown to have the ability to inhibit glycogen synthase kinase-ß(beta), which initites the wnt pathway in xenopus embryos that leads to dorsal axis formation (Klein and Melton 1996).
REFERENCES: Fredieu J., Cui Y., Maier P., Danilchik M., and Christian J. Xwnt-8 and lithium can act upon either dorsal mesodermal or neurectodermal cells to cause a loss of forebrain in Xenopus embryos. Dev. Biol. 186: 100-114, 1997. Kao K.R. and Elison R.P. The entire mesodermal mantle behaves as Spemann's organizer in dorsoanterior-enhanced Xenopus laevis embryos. Dev. Biol. 127: 64-77, 1988. Klein P.S. and Melton D.A. A molecular mechanism for the effect of lithium on development. Proc. Natl. Acad. Sci. 93: 8455-8459, 1996. Sive H. L., Grainger R.M., and Harland R.M. Early Development of Xenopus Laevis. Cold Spring Harbor: New York, 2000. Slack J.M.W., IsaacsH.V. and Darlington B.G. Inductive effects of fibroblast growth factor and lithium ion on Xenopus blastula ectoderm. Development 103: 581-590, 1988.
|
||||||||||||||||||
Last Modified: 6 April 2001
[Lab
Protocols
| Students
| Cebra-Thomas
| Course
| Links
]